

Симбиоз
5 постов
В 19 веке клир Англиканской церкви насчитывал более 15 000 человек, и сельский приходской священник, паства которого составляла примерно 250 душ, мог рассчитывать на средний доход в 500 фунтов стерлингов — столько же получали высокопоставленные чиновники.
Младшие сыновья пэров и нетитулованных мелкопоместных дворян, как правило, шли либо в церковь, либо в армию и часто вкладывали в свой приход семейное состояние. Многие имели значительный доход от сдачи в аренду церковных земель или приусадебного участка, полученного вместе с должностью.
Даже наименее привилегированные приходские священники обычно не бедствовали. Джейн Остин выросла в доме священника в Стевентоне, графство Хэмпшир, который казался ей «постыдно убогим», однако в доме были гостиная, кухня, вестибюль, кабинет, библиотека и семь спален — едва ли такое жилище можно назвать трущобой. Самый богатый церковный приход находился в Доддингтоне, графство Кембридж: он занимал 38 000 акров земли и гарантировал везучему священнику годовой доход 7300 фунтов стерлингов, что в пересчете на сегодняшние деньги составляет примерно 5 миллионов фунтов.
Священники Англиканской церкви были двух типов — викарии (vicars) и собственно священники или ректоры (rectors). Разница между ними была незначительной с точки зрения канонической и огромной — с экономической.
Церковь не платила священникам. Их доход складывался из арендной платы и церковного налога (десятины). Десятины были двух видов: «большая» десятина, которой облагались основные сельскохозяйственные культуры, такие как пшеница и ячмень, и «малая» десятина, которую платили с огородов, плодокорма (выпаса свиней в лесу) и прочих менее значительных деревенских занятий и объектов. Ректоры получали большую десятину, а викарии — малую, поэтому первые обычно были богаче вторых, порой значительно.
Десятины служили постоянным источником конфликтов между церковью и фермерами, и в 1836 году было решено упростить эту систему. Теперь вместо того, чтобы отдавать приходскому священнику оговоренную часть своего урожая, фермер платил ему фиксированную ежегодную сумму, рассчитанную исходя из общей стоимости его земли. Это означало, что духовенству полагалась точно зафиксированная выплата, и даже если у фермеров выдавался неурожайный год, у священников всегда были полные закрома.
Роль сельского священника была весьма необременительной. Набожность не являлась обязательным требованием. В Англиканской церкви рукоположение требовало университетской степени, однако большинство священников получали классическое образование и совсем не знали богословия; они не умели проповедовать, вдохновлять, утешать — словом, не могли предоставить своей пастве подлинной христианской поддержки. Многие даже не утруждали себя сочинением проповедей, а просто покупали сборники готовых проповедей и читали из книги.
В результате появился класс хорошо образованных состоятельных людей, располагавших массой свободного времени. Нередко скучающие приходские священники находили себе разные полезные хобби. Впервые в истории большая группа людей занималась целым рядом нужных дел, совершенно не входивших в их прямые обязанности.
Рассмотрим некоторые из них.
Прихожане так плохо посещали службы Джорджа Бейлдона, викария из глухого уголка Йоркшира, что он переоборудовал половину своей церкви в курятник, зато стал самоучкой-авторитетом в лингвистике и составил первый в мире словарь исландского языка.
В тех же краях, неподалеку от Йорка, приходской священник по имени Лоренс Стерн писал популярные романы, самый известный из которых — «Жизнь и мнения Тристрама Шенди, джентльмена».
![Британские [s]ученые[/s] священнослужители и наука Англия, Англиканская церковь, Наука, Наука и религия, Длиннопост](https://cs10.pikabu.ru/post_img/2019/12/22/10/1577031165182712590.jpg)
Эдмунд Картрайт, сельский священник из Лестершира, изобрел механический ткацкий станок, благодаря которому стала возможна промышленная революция. К середине 19 века в одной только Англии работали свыше 250 000 станков Картрайта.
![Британские [s]ученые[/s] священнослужители и наука Англия, Англиканская церковь, Наука, Наука и религия, Длиннопост](https://cs11.pikabu.ru/post_img/2019/12/22/10/1577031478119686926.jpg)
В Девоне преподобный Джек Рассел вывел породу терьеров, названную его именем.
![Британские [s]ученые[/s] священнослужители и наука Англия, Англиканская церковь, Наука, Наука и религия, Длиннопост](https://cs10.pikabu.ru/post_img/2019/12/22/10/1577031590172537900.jpg)
В Оксфорде преподобный Уильям Бакленд составил первое научное описание динозавров и стал ведущим в мире специалистом по копролитам — окаменелым экскрементам позвоночных.
![Британские [s]ученые[/s] священнослужители и наука Англия, Англиканская церковь, Наука, Наука и религия, Длиннопост](https://cs11.pikabu.ru/post_img/2019/12/22/10/1577031667186053879.jpg)
Томас Роберт Мальтус из Суррея написал книгу «Опыт закона развития населения» (в которой, как вы, конечно, помните, утверждается, что ускорение темпов снабжения продовольствием никогда не поспеет за ростом народонаселения), заложив тем самым начало новой дисциплины — политической экономии.
![Британские [s]ученые[/s] священнослужители и наука Англия, Англиканская церковь, Наука, Наука и религия, Длиннопост](https://cs13.pikabu.ru/post_img/2019/12/22/10/157703179719606248.jpg)
Преподобный Уильям Гринвил из Дарема был отцом-основателем современной археологии, хотя еще больше он известен среди рыбаков как изобретатель любимой многими «славы Гринвила» — искусственной мухи-наживки для ловли форели.
В Дорсете священник с необычным именем Октавиус Пикард-Кембридж стал ведущим мировым специалистом по паукам.
![Британские [s]ученые[/s] священнослужители и наука Англия, Англиканская церковь, Наука, Наука и религия, Длиннопост](https://cs12.pikabu.ru/post_img/2019/12/22/10/1577031951128572684.jpg)
Преподобный Уильям Шепард, написал историю непристойных анекдотов.
Джон Клейтон из Йоркшира устроил первую практическую демонстрацию газового освещения.
Преподобный Джордж Гарретт из Манчестера построил одну из ранних подводных лодок.
Адам Баддл, викарий-ботаник из Эссекса, вырастил цветущий кустарник, названный в его честь "буддлея".
Преподобный Джон Макензи Бейкон из Беркшира стал пионером-воздухоплавателем и отцом аэрофотографии.
Сабин Бэринг-Гулд сочинил гимн «Вперед, солдаты-христиане!» и (что гораздо более неожиданно) первый роман про оборотня — человека-волка.
Преподобный Роберт Стивен Хокер из Корнуолла писал выдающиеся стихи, которые приводили в восторг Лонгфелло и Теннисона, однако при этом несколько смущал своих прихожан тем, что носил розовую феску и почти всегда находился под умиротворяющим воздействием опиума.
Гилберт Уайт из западного Гемпшира стал самым уважаемым натуралистом своего времени, написал блестящую и до сих пор популярную книгу "Естественная история и древние памятники Селборна".
![Британские [s]ученые[/s] священнослужители и наука Англия, Англиканская церковь, Наука, Наука и религия, Длиннопост](https://cs11.pikabu.ru/post_img/2019/12/22/10/1577032477164528776.jpg)
В Нортхэмптоншире служил преподобный Майлз Джозеф Беркли, ставший выдающимся специалистом по грибам и заболеваниям растений; к сожалению, именно он оказался повинен в распространении многих вредных болезней, в том числе самой губительной для садовых культур — мучнистой росы.
![Британские [s]ученые[/s] священнослужители и наука Англия, Англиканская церковь, Наука, Наука и религия, Длиннопост](https://cs10.pikabu.ru/post_img/2019/12/22/10/1577032633153356287.jpg)
Джон Митчел изобрел метод взвешивания планеты Земля, который стал, возможно, самым оригинальным практическим научным экспериментом XVIII века.
Митчел умер раньше, чем появилась возможность осуществить этот эксперимент, который в конце концов был проведен в Лондоне Генри Кавендишем.
Но, пожалуй, наиболее необычным из всех священников был преподобный Томас Байес из Танбридж-Уэллса, графство Кент, живший в 1701–1761 годах. Этот, согласно общему мнению, робкий и совершенно безнадежный проповедник, оказался необычайно одаренным математиком. Байес вывел математическое уравнение, ныне истользующееся для решения таких сложных задач, как определение вероятностей, или обратных вероятностей, как их иногда называют. Это способ получения статистически надежных вероятностей на основе частичной информации. Самое удивительное, что до изобретения компьютера теорема Байеса не находила практического применения (поскольку только компьютер способен выполнить необходимые расчеты), так что в XVIII веке это было интересное, но бесполезное упражнение. Явно придавая мало значения своей теореме, Байес даже не потрудился ее опубликовать. В 1763 году, через два года после смерти священника-математика, один из его друзей отослал теорему в Лондонское королевское общество, где ее опубликовали в «Философских трудах» под скромным названием «Очерк о решении задачи в доктрине вероятностей». Это было ключевое событие в истории математики.
![Британские [s]ученые[/s] священнослужители и наука Англия, Англиканская церковь, Наука, Наука и религия, Длиннопост](https://cs9.pikabu.ru/post_img/2019/12/22/10/15770328211742458.png)
Сегодня теорема Байеса используется при моделировании климатических изменений, прогнозировании фондового рынка, в радиоуглеродном датировании, интерпретации космических событий и во многих других исследованиях, где необходима интерпретация вероятностей, — и все благодаря научным изысканиям английского священника XVIII века.
Многие другие представители духовенства не прославились великими делами, зато произвели на свет великих детей. Джон Драйден, Кристофер Рен, Роберт Гук, Томас Гоббс, Оливер Голдсмит, Джейн Остин, Джошуа Рейнольдс, Сэмюэл Тэйлор Кольридж, Горацио Нельсон, сестры Бронте, Альфред Теннисон, Сесиль Родз и Льюис Кэрролл (который и сам был посвящен в духовный сан, но никогда не служил) — все они были отпрысками священников.
Вся эта идиллия начала разрушаться с наступлением сельскохозяйственного упадка 1870-х, когда в течение семи лет фермеры снимали крайне низкие урожаи. В этот раз фермеры и землевладельцы не могли решить проблему простым вздуванием цен, как они обычно делали в прошлом, потому что столкнулись с сильной конкуренцией со стороны заморских производителей, особенно Америки, превратившейся в огромную сельскохозяйственную машину. Благодаря жатке Маккормика и другим грохочущим орудиям американские прерии стали устрашающе плодовиты. С 1872 по 1902 год производство пшеницы в Америке выросло на 700 %, в то время как производство пшеницы в Британии упало более чем на 40 %.
Цены тоже упали. Пшеница, ячмень, овес, бекон, свинина и баранина в последней четверти XIX века стали стоить примерно вдвое меньше, чем пару десятилетий назад. Шерсть подешевела с 28 шиллингов за четырнадцатифунтовый сверток до 12 шиллингов. Тысячи фермеров-арендаторов разорились. Более ста тысяч фермеров и рабочих ушли в города. Поля пустовали, никаких перспектив не было. Деревенские церкви стали подозрительно пустынными: прихожане разъехались кто куда, а те, кто остался, совсем обнищали. Для сельских священников настали трудные времена. Эта должность уже никогда не будет такой же привлекательной, как раньше.
На пике сельскохозяйственного кризиса британское правительство, возглавляемое либералами, сделало странную вещь. Оно ввело налог на наследство.
Изначально налог на наследство был довольно скромным (8 % за поместье стоимостью свыше одного миллиона фунтов стерлингов), но представлял собой надежный источник государственных доходов. Миллионы людей, которым не надо было его платить, поддерживали этот закон и постоянно повышали ставку налога до тех пор, пока накануне Второй мировой войны он не составил 60 % — такой уровень был разорительным даже для самых богатых землевладельцев.
В то же время государство постоянно повышало подоходный налог и придумало еще целый ряд новых налогов — налог на необрабатываемую землю, на добавленную стоимость и прочие; все они легли тяжким бременем на крупных землевладельцев. Двадцатый век стал для высших слоев общества, по словам историка Дэвида Кэннедайна, временем, когда «сгущаются тучи».
Большинство жило в состоянии перманентного кризиса. Когда случалось что-то плохое — крыша прохудилась, а налоги стали неподъемными, — исправить положение можно было только продавая фамильные ценности: картины, гобелены, драгоценности, книги, фарфор, серебряная посуда, редкие марки — все это утекало из богатых английских домов в музеи или в руки иностранцев.
Едва ли в Британии был хоть один большой дом, в котором ничего не было продано.
К началу XX века сельские священники стали получать гораздо меньше половины тех сумм, которые они получали пятьдесят лет назад. Церковный каталог Крокфорда за 1903 год уныло заявляет, что «значительная часть» духовенства «влачит жалкое существование». Далее там говорится о том, что преподобный Ф. Дж. Блисби подал 470 безуспешных заявлений на вакансию викария и в конце концов ушел жить в работный дом. Такое понятие, как «богатый священник», навсегда осталось в прошлом.
А нет финансирования - нет и науки.
Такие дела, малята...
![Британские [s]ученые[/s] священнослужители и наука Англия, Англиканская церковь, Наука, Наука и религия, Длиннопост](https://cs12.pikabu.ru/post_img/2019/05/30/6/1559209668144867595.jpg)
По книге Билла Брайсона "Краткая история быта и частной жизни".
https://www.libfox.ru/541111-112-bill-brayson-kratkaya-istor...
В очередной раз столкнулся с ситуацией: "Ты сюда машину не ставь, тут мое место".
"Владелец", парковочного места был деликатно послан, но рядом уже были парковочные места, огороженные цепочкой.
В связи с чем возник вопрос: какими документами должен обладать человек, чтобы законно претендовать на парковочное место?
Прошу прощения, если подобный пост уже был - я найти не смог.
Выжимка из книги Р. Докинза "Расширенный фенотип. Дальнее влияние гена" №11.
Спокойно! Фотографий не будет.
Паразитические и симбиотические отношения можно классифицировать очень по разному в зависимости целей классификации. Классификации, выработанные паразитологами и медиками, без сомнения эффективны в рамках их задач, но они основаны на взгляде, что хозяин - это отдельный организм, паразит - отдельный организм, и взаимодействие между ними происходит как между организмами.
Докинз же предлагает взгляд с иной стороны: есть геном хозяина, есть геном паразита, и вот они-то и являются основой взаимодействия. Исходя из этого взгляда предлагаются следующие аспекты классификации.
1. По методу выхода из хозяев и методов распространения генов хозяина и паразита.
На одном полюсе ряда будут паразиты, использующие пропагулы (пропагула -и семя, и спора, и пара особей животных разного пола, и даже одна оплодотворенная самка, т. е. все то, что впоследствии в соответствующих условиях может превратиться в популяцию) хозяина для своего собственного воспроизводства.
Такие паразиты заинтересованы в том, чтобы хозяин был жив и плодовит. И по большей части интересы хозяина совпадают с интересами паразита. Такой паразит с большой долей вероятности через какое-то время может стать симбионтом.
На другом полюсе – паразиты, гены которых распространяются не через репродуктивные пропагулы хозяина, а, к примеру – через выдыхаемый им воздух, или через мёртвое тело хозяина. В этом случае между хозяином и паразитом идёт "гонка вооружений".
2. По времени действия генов паразита в ходе развития хозяина. Ген – будь то ген хозяина или паразита, проявляет более фундаментальное влияние на конечный фенотип хозяина, если он работает на ранней фазе развития эмбриона хозяина, и менее – если включается поздно. Радикальные изменения – вроде развития двух голов, могли бы происходить в результате единственной мутации (в геноме хозяина или паразита), которая действовала бы достаточно рано в эмбриональном развитии хозяина. Позднодействующая мутация (опять же – в геноме хозяина или паразита) – мутация, которая не начинает действовать, пока тело хозяина не станет взрослым, будет вероятно, иметь лишь небольшой эффект, так как общая архитектура тела будет к тому времени уже сформирована. Поэтому паразит, который входит в своего хозяина когда тот уже взрослый – с меньшей вероятностью окажет радикальный эффект на фенотип хозяина, нежели паразит, входящий рано.
3. По "дальнодействию" генов паразита на гены хозяина.
Все гены проявляют свою силу, прежде всего – служа матрицами для синтеза белков. Поэтому локус первичной власти гена – клетка, в особенности – цитоплазма, окружающая ядро, где расположен этот ген. Потоки транспортных РНК сквозь ядерную мембрану и осуществляют генетический контроль над биохимией цитоплазмы. Тогда фенотипическая экспрессия гена – во-первых, его влияние на биохимию цитоплазмы. В свою очередь, она влияет на форму и структуру всей клетки, характер её химического и физического взаимодействия с соседними клетками. Далее этим затрагивается строение многоклеточных тканей, и в свою очередь – дифференциацию разнообразия тканей в развивающемся теле. Наконец, это проявляется в атрибутах всего организма, которые анатомы и этологи идентифицируют на их уровне – как фенотипические экспрессии генов.
Совместное влияние генов паразита и хозяина на один и тот же фенотипический признак хозяина может иметь место в любом звене только что описанной пространственной цепи. И гены улитки, и гены трематоды, паразитирующей на ней, проявляют свою власть на отдельном клеточном и даже тканевом уровне. Они влияют на химию цитоплазмы своих клеток по отдельности, потому что у них нет общих клеток. Они влияют на формирование тканей по отдельности, потому что ткани улитки не пронизаны тканями трематоды так глубоко, как например тесно пронизаны ткани водоросли и гриба в лишайниках. Гены улитки и гены трематоды влияют на развитие своих органов и систем, и более того – всех организмов по отдельности, потому что все клетки трематоды в большей степени образуют единый массив, чем нечто распределённое среди клеток улитки.
Но есть паразиты и симбионты, глубже пронизывающие ткани хозяина. Крайний случай – плазмиды и другие фрагменты ДНК, которые буквально вставляют себя в хромосомы хозяина. Более тесного паразита просто невозможно вообразить.
У вирусов есть свой белковый чехол, но они вводят свою ДНК в клетку хозяина. Поэтому они имеют возможность влиять на клеточную химию хозяина на очень низком уровне, разве что менее низком, чем уровень вставок в хозяйские хромосомы. Предполагается, что внутриклеточные паразиты в цитоплазме также могут иметь возможность проявлять значительную власть над фенотипами хозяина.
Некоторые паразиты пропитывают хозяина не на клеточном уровне, а на уровне ткани. Например – саккулина, многие паразитические грибы и растения-паразиты. При этом клетки паразита и хозяина отличны, но паразит вторгается в ткани хозяина запутанной и тонко разветвлённой корневой системой. Отдельные клетки паразитических бактерий и простейших могут пропитывать ткани хозяина столь же тесно. Такой «тканевый паразит», в слегка меньшей степени, чем клеточный, но имеет возможность сильно влиять на развитие органа, макроскопический фенотип, и на поведение. Другие внутренние паразиты, вроде только что обсуждённых трематод, не смешивают свои ткани с тканями хозяина, сохраняя их отдельно; они проявляют силу только на уровне всего организма.
Но мы еще не достигли другого полюса нашего континуума дальнодействия. Не все паразиты физически живут внутри их хозяев. Они даже могут редко входить в контакт со своими хозяевами, или не входить вовсе. Кукушка – точно такой же паразит, как и трематода. Оба – паразиты всего организма – это не тканевые и не клеточные паразиты. Если гены трематоды могут иметь фенотипическую экспрессию в теле улитки, то нет разумных причин полагать, что гены кукушки не могут иметь фенотипическую экспрессию в теле тростниковой камышовки. Различие между ними практическое, и оно явно меньше, чем различие между клеточным паразитом и тканевым. Практическое различие в том, что кукушка не живёт внутри тела тростниковой камышовки, и поэтому имеет меньше возможностей по управлению внутренней биохимией хозяина. Она должна полагаться на другие манипулятивные среды, – например звуковые или световые волны. Кукушонок использует сверхъестественно яркий зев, чтобы осуществлять управление камышовки с помощью её нервной системы, посредством её зрения. Он применяет особенно громкий просящий крик, чтобы управлять камышовкой с помощью её нервной системы через уши. Генам кукушки, проявляя своё влияние на фенотипы хозяина, приходится полагаться на дистанционное воздействие.
Предыдущие части:
https://pikabu.ru/story/znaete_li_vyi_chto_takoe_estestvenny...
https://pikabu.ru/story/moshenniki_i_modifikatoryi_6255179
https://pikabu.ru/story/_5860133
https://pikabu.ru/story/_5765793https://pikabu.ru/story/_5714779
https://pikabu.ru/story/_5710679Обиженные на бан Uspeli и его миньонов, как видно, забыли, что Пикабу не только для них создан, и решили, "не доставался же ты никому". Администрации срут, рейтинг роняют (здесь - их право, но если быть честными до конца, то ты оцениваешь приложение, а не действия его администрации), свежее завалили спамом...
Есть просьба к @moderator : баньте спамеров, а новорегам, пока страсти не улягутся - сделайте первую неделю read only.

Выжимка из книги Р. Докинза "Расширенный фенотип. Дальнее влияние гена" №10.
‒ «Знаете ли вы украинскую ночь?» ‒ с чувством начал Оська.
‒ Нет, нет!!! ‒ закричал зал. ‒ Не знаем! Просим! Просим!
‒ «Нет, вы не знаете украинской ночи!» ‒ продолжал немного смущенный Оська.
‒ Ясно, не знаем, ‒ согласились матери. ‒ Откуда нам знать? Какое наше воспитание было!
"Кондуит и Швамбрания" Л. Кассиль.
Конечно, все мы знакомы с принципами, лежащие в основе эволюции и естественного отбора:
1. Ограниченность ресурсов;
2. Выживание наиболее приспособленных;
3. Передача генетической информации потомкам;
4. Изменчивость генетической информации при копировании.
То есть, для условных сусликов, живущих в условной тундре, существуют следующие правила игры:

1. Ништяков на бесконечное количество сусликов не хватит.
2. Не все суслики доживут до взрослого возраста, а те, кто доживёт - должны ещё и с самочками замутить (а они, к слову, тоже не бессмертны).
3. Детишки будут похожи на папу и маму.
4. Но не все.
Но...

На самом деле не так все плохо. В школе вас не обманывали. Вам просто показали взгляд только с одной стороны. Со стороны индивидуального организма. И этот взгляд правильный, но не единственный.

Давайте взглянем на этих же сусликов не как на личность.

... а как на "машину для выживания", построенную вокруг ДНК суслика.

Представьте на время, что не ДНК нужна в клетке как матрица для синтеза белков, а клетка является минимальной единицей, способной реплицировать ДНК…
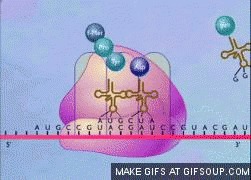
Такая концепция помещает ДНК в центр биологии.
А теперь - еще более масштабно;; на всё ДНК всех сусликов в этой тундре.
Что мы видим: вся генетическая информация многократно продублирована - своя копия есть в каждой клетке тела суслика. Там она распределена по хромосомам, но и в них порядок - каждый ген находится на своем месте - локусе.

Что же с этим массивом данных произойдет через несколько поколений?
Количество сусликов, а значит - и клеток тела, и размер самого массива останутся примерно теми же. (Мы помним, что количество ресурсов ограничено, и в условной тундре может проживать только определенное количество условных сусликов).
Структура этих данных тоже останется без изменений. В каждой клетке все столько же хромосом, а гены находятся все в тех же локусах.
Что же изменится?
Изменится соотношение аллелей - вариантов генов в одинаковых локусах. Например, если по каким-то причинам станет чуть выгоднее быть рыжим сусликом, а не серым, то выживать и плодиться рыжие будут чуть лучше серых и процент генов "рыжей окраски" возрастёт.
То есть с этой точки зрения естественный отбор работает не с организмами, а с аллелями.
Гены непосредственно конкурируют за выживание со своими аллелями, содержащимися в генофонде, поскольку эти аллели стремятся занять их место в хромосомах последующих поколений.
Организма век недолог, хромосомы в них - так же смертны - они редко могут значительно пережить организм, но гены, как информация имеющая чёткое физическое воплощение, могут существовать сотни миллионов лет без изменений.
Именно гены следует считать главной единицей отбора, и именно они играют в "Самом грандиозном шоу на Земле".
Предыдущие части;
https://pikabu.ru/story/chto_takoe_prisposoblennost_6578969
https://pikabu.ru/story/yegoistichnaya_dnk_6484677
https://pikabu.ru/story/moshenniki_i_modifikatoryi_6255179
https://pikabu.ru/story/aktivnyiy_replikator_zarodyishevoy...
https://pikabu.ru/story/manipulyatsiya_i_gonka_vooruzheniy...
https://pikabu.ru/story/ogranichenie_na_sovershenstvovanie...
https://pikabu.ru/story/vyizhimka_iz_knigi_r_dokinza_quotr...
https://pikabu.ru/story/vyizhimka_iz_knigi_r_dokinza_quotr...
https://pikabu.ru/story/vyizhimka_iz_knigi_r_dokinza_quotr...
Выжимка из книги Р. Докинза "Расширенный фенотип. Дальнее влияние гена" №9.
Предыдущие части:
https://pikabu.ru/story/yegoistichnaya_dnk_6484677
https://pikabu.ru/story/moshenniki_i_modifikatoryi_6255179
https://pikabu.ru/story/aktivnyiy_replikator_zarodyishevoy...
https://pikabu.ru/story/manipulyatsiya_i_gonka_vooruzheniy...
https://pikabu.ru/story/ogranichenie_na_sovershenstvovanie...
https://pikabu.ru/story/vyizhimka_iz_knigi_r_dokinza_quotr...
https://pikabu.ru/story/vyizhimka_iz_knigi_r_dokinza_quotr...
https://pikabu.ru/story/vyizhimka_iz_knigi_r_dokinza_quotr...
Понятие «выживание наиболее приспособленных» было введено Гербертом Спенсером в 1864 г., и было принято Дарвином в 1866 г. по настоянию Уоллеса.
В одном из писем к Дарвину, Уоллес предлагает ему уйти от использования термина «естественный отбор», поскольку этот термин часто воспринимается их оппонентами как отбор, произведенный некоей персонифицированной сущностью, Природой. В то время как спенсеровское «выживание наиболее приспособленных» лишено метафоричности и понимается однозначно.

Можно улыбнуться, мол 19 век, темнота. Однако в отношении словосочетания «эгоистичный ген» имеет место аналогичная путаница. Множество статей утверждают, что это «изобретательная, но неправдоподобная теория. Нет никаких оснований приписывать молекулам обладание такой сложной эмоцией, как эгоизм».
Ни Уоллес, ни Дарвин не могли предвидеть, что «выживание наиболее приспособленных» произведёт более серьёзную путаницу в умах, чем когда-либо производил «естественный отбор».
В первую очередь термин атаковали философы, пытаясь доказать, что теория естественного отбора является ничего не значащей тавтологией (забавный вариант – что она нефальсифицируема, а потому ложна!). Фактически же ощущение тавтологичности полностью вытекает из фразы «выживание наиболее приспособленных», а не из самой теории вообще.
Тавтологичность теории вкратце видится вот в чём: естественный отбор определяется как выживание наиболее приспособленных, а наиболее приспособленные определяются как те, кто выживает. Стало быть весь дарвинизм – нефальсифицируемая тавтология, и мы не должны далее засорять им наши головы.
Этот спор – замечательный пример возвышения слов выше их положения, который в этом отношении напоминает онтологическое доказательство существования Бога св. Ансельма. Как и Бог, естественный отбор – слишком крупная теория, чтобы её можно было бы доказать или опровергнуть, играя словами. В конце концов и Бог, и естественный отбор – единственные две имеющиеся у нас работающие теории, объясняющие наше существование.

Но даже те, кто принимает теорию эволюции, вкладывают в понятие «приспособленность» разный смысл.
1. Когда Спенсер, Уоллес и Дарвин вводили термин «приспособленность», то обвинение в тавтологии не могло быть предъявлено ни одному из них. Назовём эту первоначальную трактовку как приспособленность-1. Она не имела точного технического значения, а наиболее приспособленный определялся не как «тот, кто выживает». Приспособленность огрублённо означала способность выжить и размножиться, но она не определялась и не измерялась как нечто строго синонимичное репродуктивному успеху. Она имела диапазон специфических значений, в зависимости от конкретного аспекта жизни, исследуемого в данный момент. Если предметом внимания была эффективность размола растительной пищи, то наиболее приспособленными были особи с самыми твёрдыми зубами или наиболее мощными челюстными мускулами. В различных контекстах наиболее приспособленные особи понимались как те, кто обладал острейшим зрением, сильнейшими мускулами, острейшим слухом, быстрейшими реакциями. Предполагалось, что эти способности, наряду с бесчисленным прочими, улучшались из поколения в поколение, а естественный отбор и производил это улучшение.

«Выживание самых приспособленных» было общей характеристикой этих специфических усовершенствований. В этом нет ничего тавтологичного.
Этот смысл значения «приспособленность» - одно из самых близких к повседневному.
2. Наиболее точное и однозначное из различных технических значений – то, что принято у популяционных генетиков.
Для популяционных генетиков приспособленность – рабочий параметр, точно определяемый в рамках измерительной процедуры. Практически это слово не применяется ко всему организму, но к его генотипу, обычно в отдельном локусе. Приспособленность W генотипа, скажем Aa, может быть определена как 1−s, где s – коэффициент отбора против данного генотипа.
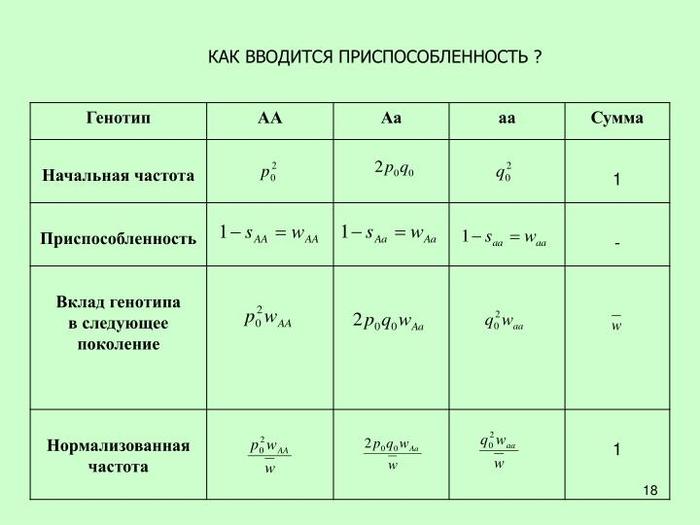
Её можно расценить как меру количества потомков, которых типичный индивидуум, обладающий генотипом Aa, вырастит до их репродуктивного возраста, в сравнении с усреднённым количеством по другим вариациям. Она обычно выражается по отношению к соответствующей приспособленности одного конкретного генотипа в локусе, который произвольно определен как 1. Тогда говорят об отборе в том локусе в пользу генотипов с более высокой приспособленностью, по отношению к генотипам с более низкой приспособленностью. Назовём это специальное значение термина популяционных генетиков приспособленностью-2. Когда мы говорим, что кареглазые особи приспособленней голубоглазых, то мы говорим о приспособленности-2. Мы предполагаем, что все другие вариации у особей усреднены, и мы фактически применяем слово «приспособленность» к двум генотипам в отдельном локусе.
3. Пока популяционные генетики интересуются непосредственными изменениях частот генотипа и частот гена, этологи и экологи рассматривают целые организмы как интегрированные системы, которые кажется что-то максимизируют. Приспособленность-3, или «классическая приспособленность», является свойством индивидуального организма, часто выражаемым как результат выживания и плодородия. Она часто измеряется как количество его потомков, выращенных до взрослого состояния, но имеют место определённые споры относительно полезности этого.

Если просто считать количество рожденных детей, то не учитывается детская смертность. Если она оценивается как количество детей, достигших репродуктивного возраста, то она не будет учитывать изменения в репродуктивном успехе этих детей. Если она оценивается как количество внуков, то она не учитывает…и так далее до бесконечности. В идеале мы можем подсчитать относительное количество живых потомков спустя какое-то, очень большое количество поколений. Но такая «идеальная» оценка имеет любопытную особенность – если довести её до логического конца, то она может принимать только два значения: всё – или ничего. Если мы заглянем в будущее достаточно далеко, то мы обнаружим только две альтернативы: или у нас вообще не будет потомков, или все живые люди будут нашими потомками.
Это несовершенство термина «приспособленность-3» привело к появлению новых его трактовок.
4. Гамильтон учел, что естественный отбор одобрит органы и поведение, которые позволяют генам индивидуума передаваться независимо от того, является ли этот индивидуум непосредственным предком, или нет. Индивиды, помогающие своему брату быть предком, может этим самым гарантировать выживание в генофонде генов «братской поддержки». Гамильтон увидел, что родительская забота – лишь частный случай заботы о близких родственниках, с высокой вероятностью содержащих гены такой заботы. Классическая приспособленность-3 – репродуктивный успех, была слишком узка. Её надлежало расширить до «итоговой приспособленности», которая будет здесь называться приспособленностью-4.
Итоговая приспособленность индивида – это его собственная приспособленность-3 плюс по половине приспособленности-3 каждого его брата (гены братьев совпадают на 50%) плюс одна восьмая приспособленности-3 каждого кузена, и т.д
Но этот подход позволяет подсчитывать детей много раз, словно у них много параллельных жизней. Когда рождается ребёнок, дополняя уже имеющееся множество братьев, то итоговая приспособленность всех ранее рождённых братьев, согласно этому представлению, немедленно поднимется на равную величину, независимо от того, пошевелил ли кто-то из них пальцем для кормления младенца. Итоговая приспособленность другого, ещё нерожденного брата теоретически увеличилась бы при рождении его старшего племянника. Более того, этот более поздний брат мог бы быть абортирован вскоре после зачатия и тем не менее, согласно этому ошибочному представлению, наслаждаться веской «итоговой приспособленностью» благодаря потомкам его старших братьев. Если довести эту редукцию до абсурда, то он даже не обязан утруждать себя зачатием, чтобы тем не менее иметь высокую «итоговую приспособленность»!
Гамильтон ясно видел этот софизм, и потому его концепция итоговой приспособленности была более изощрённей. Итоговая приспособленность организма – не есть его свойство, но свойство его деятельности или влияний. Итоговая приспособленность подсчитывалась как собственный репродуктивный успех индивидуума плюс его влияние на репродуктивный успех его родственников, взятый с соответствующим коэффициентом родства. Поэтому, если например мой брат эмигрирует в Австралию, и я после этого никак не могу повлиять на его репродуктивный успех, то моя итоговая приспособленность не повышается при каждом рождении им ребёнка.
Чтобы определить влияние индивидуума на выживание и воспроизводство его родственников, мы должны сравнивать влияние совокупности всех поступков его жизни, и сравнить их с гипотетической жизнью в полном бездействии, словно он никогда не был зачат. Именно это последнее понимание обычно обозначает итоговую приспособленность индивидуального организма.
5. Приспособленность-5, можно трактовать как своего рода итоговую приспособленность, поставленную задом наперёд. Тогда как итоговая приспособленность-4 фокусируется на влиянии изучаемого индивида на приспособленность-3 его родственников, персональная приспособленность-5 фокусируется на влиянии родственников индивидуума на его приспособленность-3. Приспособленность-3 индивидуума – некоторая мера количества его детей или более далёких потомков. Но Гамильтон показал нам, что индивидуум может породить больше потомков, чем он мог бы вырастить сам, потому что его родственники вносят вклад в выращивание части его потомства.
Практическое преимущество использования персональной приспособленности-5, а не итоговой приспособленности-4 состоит в том, что мы прекращаем просто считать потомков, и нет никакого риска по ошибке подсчитать данного ребенка много раз.
Хотя приведенный список из пяти пунктов уже достаточно запутан, это далеко не все возможные трактовки. Например, «по причине заинтересованности в биологическом «прогрессе»», Тоди предлагает «приспособленность долгосрочного развития», определяемую как вероятность того, что линия продолжится в течение очень долгого времени, таком как 108 поколений.
Заметьте, что все эти приспособленности, кроме приспособленности-2, крепко привязаны к представлению об индивидуальном организме как «максимизирующем агенте». Докинз шутит, что приспособленность индивидуального организма - это «свойство индивидуального организма, которое будет казаться максимизирующимся тогда, когда на деле максимизируется выживание гена», и смотрит на это понятие, как на последний рубеж обороны, последнюю попытку спасти индивидуальный организм как уровень, на котором мы рассматриваем естественный отбор.
Выжимка из книги Р. Докинза "Расширенный фенотип. Дальнее влияние гена" №8.
Предыдущие части:
https://pikabu.ru/story/moshenniki_i_modifikatoryi_6255179
https://pikabu.ru/story/aktivnyiy_replikator_zarodyishevoy...
https://pikabu.ru/story/manipulyatsiya_i_gonka_vooruzheniy...
https://pikabu.ru/story/ogranichenie_na_sovershenstvovanie...
https://pikabu.ru/story/vyizhimka_iz_knigi_r_dokinza_quotr...
https://pikabu.ru/story/vyizhimka_iz_knigi_r_dokinza_quotr...
https://pikabu.ru/story/vyizhimka_iz_knigi_r_dokinza_quotr...
Теперь обратимся к другому крупному классу мошенников - горизонтальным мошенникам. Это мошенники настолько непослушные, что уходят в целом от соревнования аллелей в границах локуса. Они распространяются на другие локусы, даже создают новые локусы для себя, увеличивая размер генома. В "Nature" их окрестили "Эгоистичная ДНК".

Явно излишняя ДНК может быть в различной степени повторяющейся или бессмысленной в терминах генетического кода. Какие-то её части вероятно никогда не транскрибируются в РНК. Другие могут быть транскрибированы в РНК, но затем "вырезаны" до того, как РНК транслируется в последовательность аминокислот. Так или иначе, но она никак не проявляет себя фенотипически.

Биологи изнуряют свои умы, стараясь понять, что же этот излишек ДНК делает. Для неё были предложены различные функции.
Например, что наличие некодирующей ДНК раздвигает гены в пространстве, что увеличивает вероятность разделения их при кроссинговере и уменьшает количество связанных генов. Или что большой объем ДНК нужен для большого размера клеточного ядра.
Но все эти гипотезы основаны на идее о том, что ДНК, как и любая другая часть организма, отобрана отбором потому, что делает организму некое благо. Гипотеза эгоистичной ДНК базируется на обратном предположении: фенотипические признаки имеют место потому что они помогают ДНК копировать себя, и если ДНК сможет найти более быстрые и лёгкие способы самокопирования без синтеза белка - она будет отобрана отбором именно за такое поведение.
Живая клетка, особенно ядро эукариотов, напичкана активными механизмами репликации и рекомбинации нуклеиновых кислот. ДНК-полимераза с готовностью катализирует репликацию любой ДНК; ей безразлично, является ли данная ДНК значащей в терминах генетического кода. "Вырезание" фрагментов ДНК, и "вклейка" в неё других фрагментов ДНК - это всё части нормального инструментария в деятельности клеточного аппарата, ибо они происходят каждый раз, когда случается кроссинговер или другой тип рекомбинации Фрагменты ДНК могут быть вырезаны из одной части генома, и вставлены в другую с небрежной лёгкостью.
А вот здесь на них можно посмотреть:
https://pikabu.ru/story/vashi_udivitelnyie_molekulyarnyie_ma...

Репликабельность и "вклеиваемость", представляются одними из наиболее существенных свойств ДНК в её естественной окружающей среде клеточных машин.
Учитывая возможности такой окружающей среды, учитывая существование клеточных фабрик, существующих для репликации и склейки ДНК, было бы естественно ожидать, что естественный отбор одобрит варианты ДНК, способные эксплуатировать эту ситуацию ради своей выгоды. Выгода в данном случае означает просто многократную репликацию в зародышевых линиях. Любая вариация ДНК, свойства которой по случаю делают её с готовностью копируемой, автоматически станет распространённейшей в мире.
Как это ни парадоксально, но мы более всего знакомы с косвенными, сложными, и окольными методами, которыми молекулы ДНК обеспечивают своё будущее. Это их эффекты воздействия на тела (фенотипические эффекты), достигаемые путём управления синтезом белка, и следовательно - периферическим маршрутом управления эмбриональным развитием, непосредственно влияют на морфологию, физиологию и поведение особи. Но существуют также и гораздо более прямолинейные и простые пути, которыми варианты ДНК могут распространяться за счёт конкурирующих вариантов (аллелей).

Обычный отбор приводит к изменениями частот репликаторов по отношению к их аллелям в определенных локусах хромосом популяций. Внутригеномный отбор эгоистичной ДНК - иной вид отбора. Здесь не имеет места относительный успех аллелей ни в одном локусе генофонда, но значима способность к распространению некоторых видов ДНК на различные локусы или к созданию новых локусов. Кроме того, отбор эгоистичной ДНК не привязан к смене поколений особи; он может селективно возрастать при любом митотическом делении клетки в зародышевых линиях развивающихся тел.
В каком смысле эгоистичная ДНК является мошенником? В таком, что организмы выиграли бы от её отсутствия. Возможно она вредит, впустую занимая место и потребляя молекулярное сырьё, возможно - требует расточительного расхода ценного "машинного времени" механизмов коррекции и дублирования. В любом случае, мы можем ожидать, что отбор будет стремиться устранить эгоистичную ДНК из генома. А пока в огромных "машинах для выживания" вместе с "обычными" генами путешествует также множество хоть и безвредных, но и бесполезных реплицирующихся попутчиков, которые известны под различными именами, в зависимости от размера и свойств: плазмиды, эписомы, мобильные элементы, плазмоны, вирионы, транспозоны, репликоны, вирусы.

Выжимка из книги Р. Докинза "Расширенный фенотип. Дальнее влияние гена" №7.
Предыдущие части:
https://pikabu.ru/story/aktivnyiy_replikator_zarodyishevoy_l...
https://pikabu.ru/story/manipulyatsiya_i_gonka_vooruzheniy_v...
https://pikabu.ru/story/ogranichenie_na_sovershenstvovanie_v...
https://pikabu.ru/story/vyizhimka_iz_knigi_r_dokinza_quotras...
https://pikabu.ru/story/vyizhimka_iz_knigi_r_dokinza_quotras...
https://pikabu.ru/story/vyizhimka_iz_knigi_r_dokinza_quotras...
Естественный отбор – это процесс, с помощью которого репликаторы взаимораспространяют друг друга. Они делают это, оказывая фенотипические эффекты на мир, и часто удобно рассматривать эти фенотипические эффекты сгруппированными вместе в дискретные «носители» - индивидуальные организмы.

Этот взгляд создаёт питательную почву для мнения, что естественные отбор отбирает гены, повышающие приспособленность организма. Ген помогает выживать организму, организм размножается, создает копии полезного гена, а заодно - и других генов в геноме организма. То есть, существует некое «сотрудничество» между генами в разных локусах.

Обычно, так оно и есть.
Но также легко представить себе ситуации, в которых ген мог бы поддерживать своё собственное выживание, нанося вред возможностям выживания большей части остального генома. Такие гены называются мошенниками.
Докинз пишет о двух главных классах мошенников.

«Аллельный мошенник» – определяется как репликатор, имеющий положительный коэффициент отбора в своём локусе, но в большинстве других локусов действует отбор в сторону подавления его эффекта (в его локусе). Примеры – «гены-нарушители расщепления» или гены «мейотического драйва». Они одобряются в своих локусах благодаря тому, что попадают более чем в 50% произведённых гамет.
Особенно хорошо изучены SD-гены (SD – Segregation distorter – нарушители расщепления) у дрозофилы, которые при мейозе «ломают» сперматозоид, получающий нормальную хромосому. Пока гомологичные хромосомы всё ещё разделены на пары в ходе мейоза, SD-хромосома может проделать нечто над её нормальным партнёром (и конкурентом) такое, что позже вызовет дисфункцию сперматозоида.
Возникает следующая ситуация: если организм гомозиготен по SD-хромосоме, то она будет передана его потомкам. Если же он гетерозиготен, то при мейозе SD-гены разных хромосом ломают друг другу сперматозоиды и организм становится бесплодным. Генофонд в целом страдает, но SD-гены распространяются хорошо.
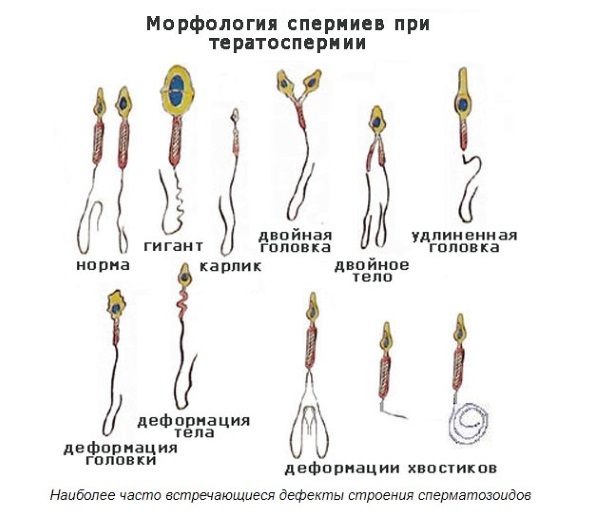
А нарушитель расщепления на половой хромосоме – это просто бомба. Он не просто создает вредные побочные эффекты, он искажает соотношение полов, и даже может совсем исключить один пол из всей популяции. Гамильтон смоделировал на компьютере популяцию, состоящую из 1000 самцов и 1000 самок, в которую был подставлен единственный мутантный самец с «преимущественно переносимой» Y-хромосомой, вынуждающей самцов иметь только сыновей, но никак не дочерей. Потребовалось только 15 поколений, чтобы моделируемая популяция вымерла из-за отсутствия самок.

В таких условиях гены в других локусах, эффект которых заключается в подавлении искажения расщепления, будут одобрены отбором. Подавить исказителя расщепления - единственный способ не окончить свой жизненный путь в бесплодном теле. Такие гены получили название «модифкаторы».

Возникает гонка вооружений между генами – аллельными мошенниками и генами – модификаторами. Перевес почти всегда на стороне модификаторов, так как они могут возникнуть в множестве локусов и сообща подавить вредный фенотипический эффект мошенника.
А вот тут начинается веселье:
- Модификаторы сами могут приводить к вредным фенотипичским эффектам. Однако, лучше с ними, чем без них.
- На стороне мошенника может оказаться сцепленный с ним ген и начать борьбу с модификаторами (своими фенотипическими эффектами стараться подавить эффекты модификаторов).
- Если в геноме окажется два разнородных мошенника, то они будут мешать друг другу.
- Наконец, через несколько поколений, основной фенотипический эффект гена-расщепителя подавлен. Но в результате геном битком набит генами-супрессорами. А это тоже можно рассматривать как эффект гена-мошенника.

Небольшое отступление, которое нужно читать очень медленно. Я бы вынес его в отдельную часть, но, боюсь, что она будет не понята теми, кто не прочитал про модификаторы.
Доминантность и рецессивность – свойства не самих генов, а свойств их фенотипических эффектов. Более того - выгодные эффекты данного гена постепенно становились доминирующими благодаря отбору модификаторов, а вредные эффекты – постепенно становились рецессивными. В самом деле – некий ген может быть доминирующим по одному из его плейотропных эффектов, и рецессивным по другому.

Фенотипический эффект гена – есть совместный продукт его самого, и окружающей его среды, включающей и другие гены в геноме.
Хотя эти «другие гены» известны как модификаторы, сейчас стало понятно, что не существует отдельной категории генов-модификаторов, отличающихся от главных генов. Вернее, любой ген может служить модификатором фенотипических эффектов любого другого гена. Да, фенотипические эффекты любого гена восприимчивы к модификации многими другими генами в геноме, генами, которые сами могут иметь много других – как главных, так и вспомогательных эффектов.

Другой крупный класс мошенников -«горизонтально распространяющиеся гены» – мы рассмотрим в следующей части.



